






Nanofert anuncia novo CEO e aposta em crescimento no Brasil
Nova gestão impulsiona expansão de nanofertilizantes para agricultura sustentável
11.08.2025 | 17:17 (UTC -3)
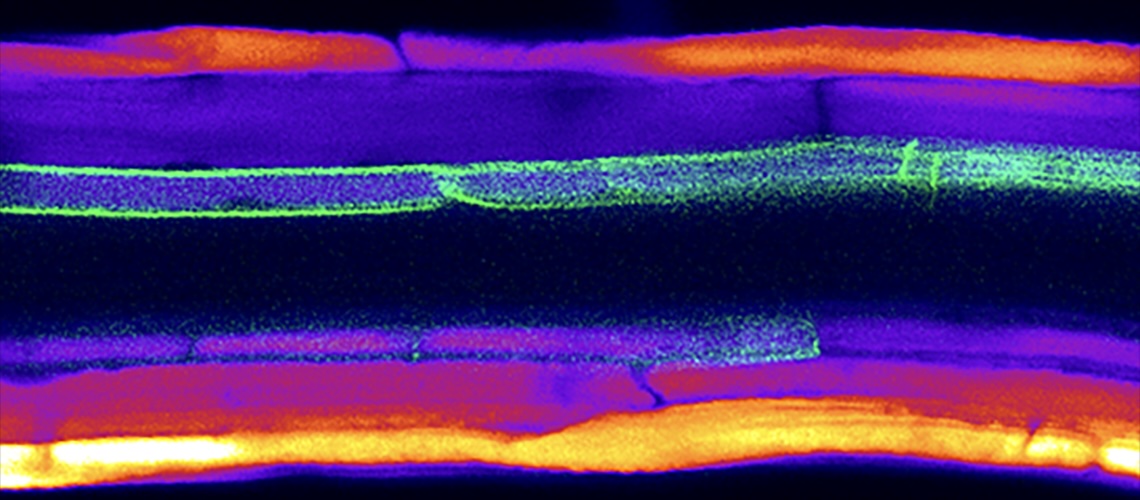
Transporte célula a célula persiste em raízes maduras mesmo após o desenvolvimento completo das barreiras apoplásticas. Essa é a principal constatação de um estudo conduzido por pesquisadores da Universidade de Genebra.
Os cientistas demonstraram que os plasmodesmata — canais citoplasmáticos que conectam células vegetais adjacentes — continuam ativos em raízes de Arabidopsis thaliana já diferenciadas. Mesmo após a formação das faixas de Caspary e da deposição de lamelas de suberina na endoderme, o transporte de moléculas do epiderma até o periciclo continua ocorrendo. O mecanismo depende dos plasmodesmata e segue um padrão unidirecional, orientado para o interior da raiz.
A pesquisa utilizou marcadores fluorescentes e linhas transgênicas com expressão de GFP (proteína verde fluorescente) sob controle de promotores específicos. As observações foram feitas por microscopia de fluorescência e por análises quantitativas de intensidade de sinal.
As raízes jovens permitem o transporte bidirecional de moléculas entre todas as camadas celulares. À medida que amadurecem, porém, o transporte se torna unidirecional, do epiderma para o centro da raiz. Esse padrão sugere um controle ativo do fluxo, contrário à ideia anterior de difusão passiva em todas as direções.
A direção do fluxo não depende apenas da presença de plasmodesmata. Os pesquisadores testaram mutantes deficientes nas faixas de Caspary (sgn3) e na suberina (ELTP::CDEF1), sem observar alteração na direção do transporte. A direção também não pôde ser explicada por deposição de calose, substância que fecha plasmodesmata. Análises com anticorpos e corantes específicos mostraram ausência de calose nas zonas diferenciadas.
A força motriz por trás do transporte foi atribuída ao fluxo de água. Ao aplicar sorbitol, um agente osmótico, os pesquisadores observaram inibição do movimento de GFP entre camadas celulares. A redução no fluxo hídrico impediu o transporte, sem induzir deposição de calose. Isso indica que o movimento de moléculas entre células ocorre por arraste hidrodinâmico, sustentado por gradientes de pressão gerados pelo sistema vascular da planta.
Para investigar os mecanismos que determinam essa direção, os autores realizaram uma triagem genética e identificaram o mutante rol1/ssm1, que rompe a barreira de unidirecionalidade. Esse mutante exibe transporte célula a célula em ambas as direções, mesmo em raízes completamente diferenciadas.
A mutação afeta o gene ROL1/SSM1, que codifica uma sintase de UDP-L-ramnose. A ramnose é essencial na síntese de pectinas do tipo rhamnogalacturonana-I e II, componentes fundamentais da parede celular. O mutante apresenta alterações globais na composição e organização da parede celular, especialmente em pectinas e celulose. A deposição de pectina torna-se desorganizada, com sinais acumulados nos cantos das células e redução entre paredes adjacentes.
As paredes celulares do mutante apresentam maior porosidade e arquitetura anômala. Imagens de microscopia eletrônica revelaram plasmodesmata mais largos em comparação com o tipo selvagem. A largura aumentada — cerca de 57% — eleva o limite de exclusão de tamanho dos plasmodesmata. Moléculas maiores, como versões duplicadas de GFP (2xGFP), atravessaram as barreiras intercelulares em rol1/ssm1, mas não no tipo selvagem.
O mutante também demonstrou melhor desempenho sob estresse hídrico. Em testes com sorbitol e em condições de seca em solo, as plantas rol1/ssm1 mostraram maior tolerância, com recuperação superior após reidratação. Essas plantas, apesar de menores e com raízes mais curtas, apresentaram padrões de crescimento normais e fertilidade preservada.
A análise iônica revelou apenas alterações sutis: redução nos teores de ferro e cálcio nas raízes, menor translocação de cálcio para os caules e aumento da transferência de cobre. A restrição na mobilização de cálcio parece refletir a necessidade de sua concentração nas camadas internas da raiz antes do carregamento no xilema, etapa considerada limitante para esse nutriente.
Outros mutantes com alterações na parede celular, como mur1, qua2, lrx1, fer, rao1/cdk8 e tt4, não apresentaram o mesmo fenótipo de transporte simétrico observado em rol1/ssm1. A exceção ocorreu apenas nas duplas combinações desses mutantes com rol1, sugerindo que o fenótipo “abre-te sésamo” é específico da perda de função de ROL1/SSM1.
Apesar do aumento da conectividade entre células, o mutante manteve seletividade de tamanho nos plasmodesmata. Proteínas GFP triplas (3xGFP), com massa de 81 kDa, não conseguiram atravessar os canais, permanecendo retidas na endoderme. Isso indica que, embora o tamanho dos poros tenha aumentado, os plasmodesmata de rol1/ssm1 não se tornaram completamente permeáveis.
O estudo também propõe um modelo onde a combinação entre estrutura dos plasmodesmata e fluxo de água define a direcionalidade do transporte celular. Alterações estruturais nos canais, como observado em rol1/ssm1, podem neutralizar a polaridade, favorecendo o fluxo em ambas as direções. Essa hipótese reforça a ideia de que o controle do transporte entre células é dinâmico e regulado por múltiplos fatores, incluindo composição da parede celular, arquitetura dos canais e forças físicas locais.
Os dados obtidos revelam o papel funcional da pectina na morfologia e na permeabilidade dos plasmodesmata. Defeitos em sua biossíntese provocaram mudanças no transporte e na arquitetura celular. As observações sugerem que a estrutura das pectinas — especialmente os polímeros do tipo RG-I — influencia diretamente a conformação dos canais intercelulares.
Plasmodesmata com abertura aumentada podem facilitar a comunicação celular, mas também introduzem riscos de perda de seletividade. A persistência da seletividade em rol1/ssm1 indica que os mecanismos de controle permanecem, ainda que modificados.
Outras informações em doi.org/10.1016/j.molp.2025.07.004
Receba por e-mail as últimas notícias sobre agricultura


Receba por e-mail as últimas notícias sobre agricultura



