


Pulgões do trigo desafiam resistência genética
Revisão científica aponta papel da saliva dos insetos e defesas em camadas no desenvolvimento de cultivares resistentes
28.06.2026 | 15:39 (UTC -3)
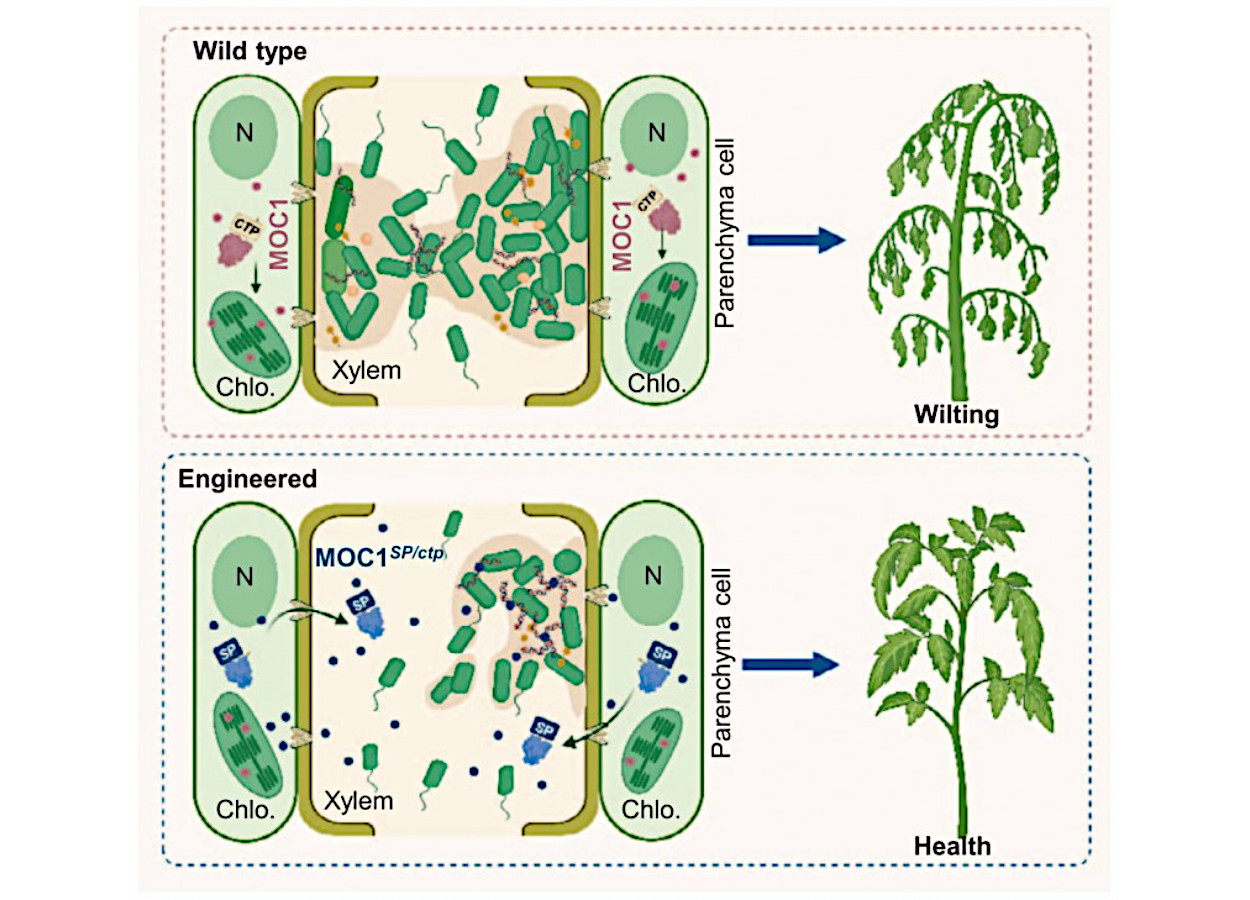
Pesquisadores chineses desenvolveram estratégia genética para aumentar a resistência de tomate e arroz contra doenças bacterianas vasculares. O trabalho reposicionou a enzima vegetal MOC1, antes direcionada ao cloroplasto, para o apoplasto. Com essa mudança, a proteína passou a atuar perto dos biofilmes formados por bactérias no xilema (DOI 10.1093/hr/uhag046).
A proposta mira uma limitação comum no controle de patógenos vasculares. Bactérias como Ralstonia solanacearum e Xanthomonas oryzae pv. oryzae invadem vasos do xilema, avançam pelo sistema vascular e prejudicam o transporte de água. Essas bactérias formam biofilmes, estruturas capazes de proteger as células bacterianas contra defesas da planta e favorecer a permanência nos tecidos.
O alvo escolhido pelos pesquisadores recebeu atenção por causa do papel do DNA extracelular, ou eDNA, na estabilidade do biofilme. Esse material funciona como uma base estrutural da comunidade bacteriana. A equipe explorou a atividade de MOC1, uma resolvase de junções de Holliday conservada em plantas, capaz de agir sobre estruturas de DNA. Em sua forma natural, MOC1 segue para o cloroplasto. Por isso, não alcança os biofilmes bacterianos no xilema.
Para resolver esse problema de localização, os cientistas substituíram o peptídeo de trânsito ao cloroplasto por um peptídeo sinal de secreção. No tomate, a equipe usou o sinal de secreção da proteína relacionada à patogênese SlPR1. A versão modificada de SlMOC1 acumulou no apoplasto. A SlMOC1 recombinante também reduziu o eDNA liberado por Ralstonia solanacearum e desorganizou a rede de eDNA presente no biofilme bacteriano.
Em tomateiro da cultivar Moneymaker, plantas transgênicas com SlMOC1 secretada apresentaram sintomas mais leves de murcha bacteriana após inoculação por encharcamento do solo com a cepa GMI1000 de Ralstonia solanacearum. As plantas também tiveram menor índice de doença e menor movimentação bacteriana nos vasos, em comparação com plantas selvagens.
O ganho de resistência não decorreu de ativação ampla da imunidade vegetal. Os pesquisadores avaliaram genes de defesa no tomate e não observaram indução de SlPR1, SlERF2a e SlEDR1 nas plantas com SlMOC1 secretada. Esse resultado indica ação ligada, principalmente, à ruptura física do biofilme bacteriano.
A equipe também testou controle mais preciso da expressão. Para isso, usou o promotor SlERF2b, induzido por infecção de Ralstonia solanacearum, para dirigir a produção de SlMOC1 secretada. As plantas mostraram atraso na murcha, menor incidência de doença aos quatorze dias após a inoculação e menor bloqueio vascular. Amostras de xilema das plantas modificadas apresentaram poucas bactérias aderidas e menor formação de biofilme.
No arroz, os cientistas aplicaram lógica semelhante. A equipe inseriu OsMOC1 com peptídeo de secreção de OsPR1b sob controle do promotor constitutivo Ubiquitin 1, no fundo genético ZH11. Após inoculação por corte de folha, em condições de campo, as plantas modificadas apresentaram lesões menores causadas por três cepas de Xanthomonas oryzae pv. oryzae: YN11, FuJ23 e PXO99A.
Os pesquisadores também desenvolveram um promotor sintético induzido por efetores TALE de Xanthomonas. O arranjo reuniu elementos de ligação a efetores, ou EBEs, associados a dez TALEs conservados em cepas asiáticas de Xanthomonas oryzae pv. oryzae. Em arroz, esse sistema ativou OsMOC1 secretada após infecção por PXO99A. As plantas EBEXoo::OsMOC1SP/ctp tiveram redução superior a sessenta e cinco por cento no comprimento das lesões, em comparação com ZH11.
A microscopia eletrônica de varredura reforçou o efeito sobre o biofilme. Em folhas selvagens, vasos do xilema apareceram preenchidos por bactérias envolvidas por matriz espessa. Nas plantas com expressão induzida por EBEs, os vasos continham poucos agregados bacterianos e biofilme mais fino.
O estudo também avaliou possíveis custos ao desenvolvimento. Em tomate, plantas com expressão induzida por SlERF2b mantiveram altura e peso de frutos comparáveis aos do tipo selvagem. Em arroz, as linhagens modificadas não apresentaram diferenças significativas em teor relativo de clorofila, altura de planta, comprimento da folha bandeira, número de perfilhos, desenvolvimento de panícula ou peso de sementes, conforme os ensaios descritos no artigo.
O modelo proposto pelos pesquisadores descreve uma mudança de destino celular da enzima. Em plantas selvagens, MOC1 segue para o cloroplasto e não entra em contato com o biofilme no xilema. Na planta modificada, a troca do peptídeo de trânsito por um sinal de secreção envia MOC1 ao espaço extracelular. Ali, a enzima rompe a rede de eDNA do biofilme, reduz a colonização bacteriana e aumenta a resistência vascular.
Os pesquisadores indicam a relocalização cloroplasto-apoplasto como uma plataforma para melhoramento de resistência a doenças vasculares dependentes de biofilme. Eles também apontam o uso futuro de edição por CRISPR para substituir, no próprio genoma, sequências nativas de direcionamento ao cloroplasto por sinais de secreção. Essa abordagem poderia gerar germoplasma editado sem DNA transgênico e com maior imunidade vascular.
Receba por e-mail as últimas notícias sobre agricultura


Receba por e-mail as últimas notícias sobre agricultura



