






Primeira detecção de "Dalbulus maidis" em Oklahoma gera preocupação
Há alerta para possível expansão da praga para outras áreas do cinturão de milho dos Estados Unidos
08.10.2024 | 18:00 (UTC -3)
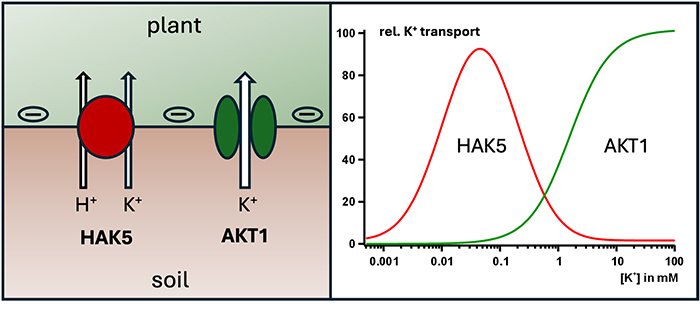
Pesquisas recentes demonstram que o transportador HAK5 desempenha papel crucial na absorção de potássio em solos pobres. O potássio é um macronutriente essencial. Quando há baixa concentração desse elemento no solo, a planta depende de mecanismos que permitam sua absorção eficiente em condições adversas.
O canal iônico AKT1 e o transportador HAK5 cooperam na captação de potássio pelas raízes. Em solos com maior concentração de potássio, o AKT1 é capaz de realizar o transporte sem grandes custos energéticos. No entanto, quando o nível de potássio cai para valores muito baixos, o HAK5 assume a função de absorção, necessitando, além do campo elétrico, de um gradiente de pH. Esse gradiente é essencial para que o HAK5 funcione e custa energia à planta.
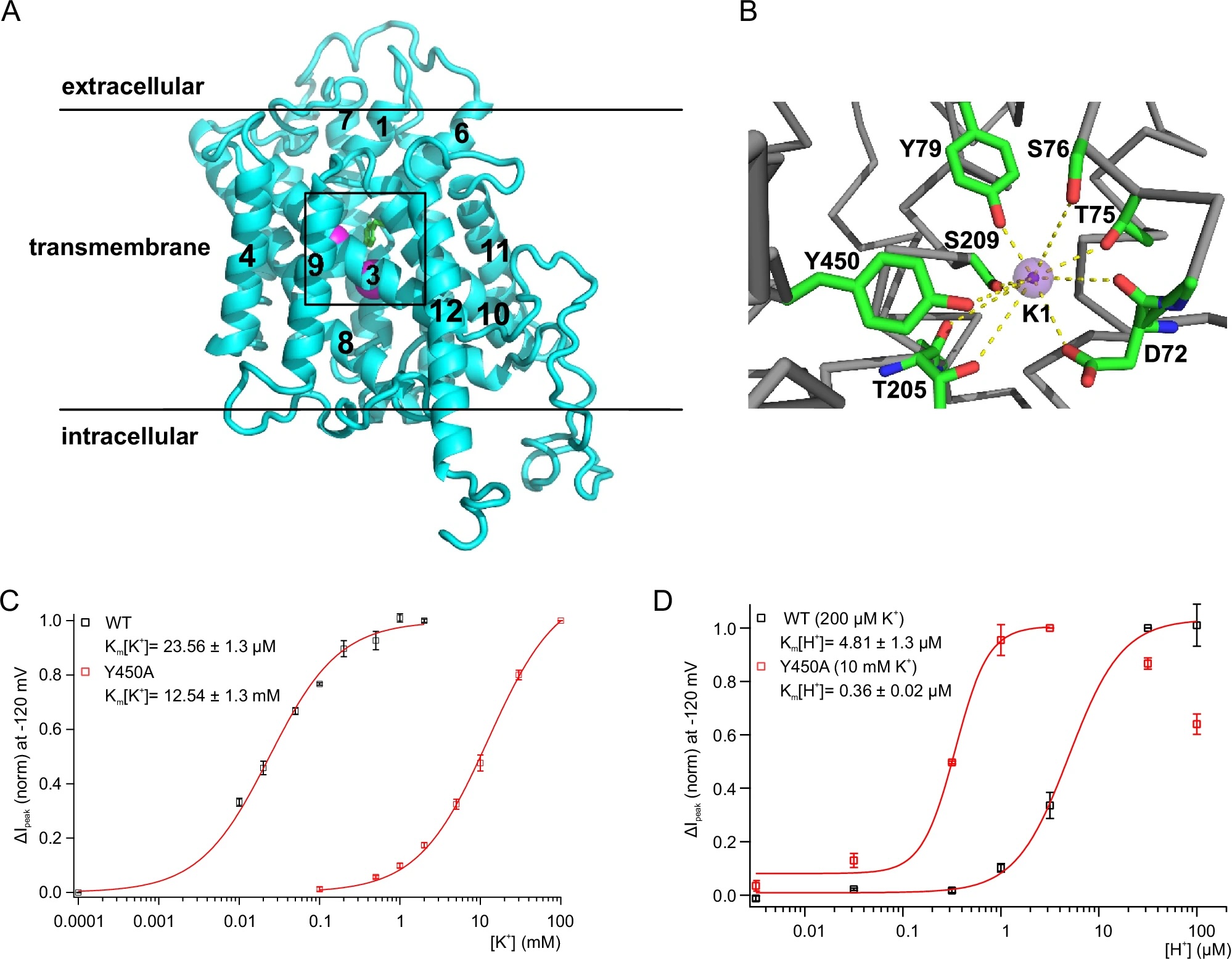
Estudos conduzidos por uma equipe da Universidade de Würzburg, liderada pelo professor Rainer Hedrich, revelaram que, enquanto o AKT1 depende de maiores concentrações de potássio para operar, o HAK5 funciona mesmo em níveis muito baixos. A pesquisa avançou no entendimento de como o transportador HAK5 é ativado, mostrando que ele possui um sensor de potássio, crucial para detectar a quantidade disponível no solo.
A compreensão do mecanismo de transporte do HAK5 pode ser revolucionária para a agricultura. Solos pobres em potássio exigem grandes quantidades de fertilizantes. Isso representa custo elevado para os produtores. A modificação genética de culturas para melhorar a eficiência do uso de potássio pode reduzir essa dependência, promovendo uma agricultura mais sustentável.
A equipe também identificou mutação que reduz drasticamente a afinidade do HAK5 pelo potássio, sugerindo que a regulação desse transportador é mais complexa do que se imaginava. Essa descoberta pode ser aplicada para melhorar a tolerância das plantas a solos com baixos níveis de potássio, o que é de extrema importância em regiões com solos pobres.
Outro ponto importante revelado pelos pesquisadores é a cooperação entre o canal AKT1 e o transportador HAK5. O AKT1 opera eficientemente em solos com níveis médios e altos de potássio. Mas, quando o nível de potássio no solo cai, o HAK5 entra em ação, garantindo que a planta continue a absorver potássio de forma eficiente.
Esses dois mecanismos são ativados de acordo com a necessidade da planta e a disponibilidade de potássio no ambiente. Essa adaptação permite que as plantas sobrevivam em solos pobres em nutrientes, o que pode ser essencial para o cultivo em áreas menos férteis.
Embora os avanços no entendimento do HAK5 sejam promissores, muitos aspectos de sua regulação ainda precisam ser explorados. Rainer Hedrich e sua equipe planejam investigar como o transporte de potássio para dentro da célula radicular está acoplado ao transporte de prótons. Entender essa relação pode abrir novas portas para a engenharia genética de plantas mais resistentes e com maior eficiência na utilização de potássio.
Mais informações podem ser obtidas em doi.org/10.1038/s41467-024-52963-6
Receba por e-mail as últimas notícias sobre agricultura


Receba por e-mail as últimas notícias sobre agricultura



